Naše skupina se v současné době zaměřuje na tři provázaná témata:
- Biogeneze a kontrola kvality fotosystémů.
- Úloha sinicových homologů bakteriálních FtsH proteáz ve fotosyntéze a dalších buněčných procesech.
- Modifikace NtcA regulonu zajišťujícího souhru metabolismu dusíku a uhlíku a možnosti jeho využití v biotechnologii.
Jako experimetální model používáme glukózo-tolerantní kmen sinice Synechocystis PCC 6803, který s vyznačuje rychlým růstem, přirozenou transformabilitou a možností kultivace na glukóze, pokud jsou vyřazeny geny nezbytné pro fotosyntézu. Díky těmto vlastnostem je Synechocystis vynikajícím modelem pro studium základních procesů na fotosyntetických membránách společných pro sinice, řasy i rostliny. Využíváme moderní metody molekulární biologie, biochemie proteinů a zobrazování buněk, a ve spolupráci s partnery i pokročilou strukturní biologii.
Náš výzkum je v současné době podporován projekty uvedenými zde a zde.
Biogeneze a kontrola kvality fotosystémů
Fotosystémy I a II (PSI a PSII) jsou zodpovědné za kyslíkovou fotosyntézu, biochemický proces nezbytný pro evoluční vývoj a udržení života na Zemi. Na těchto komplexech dochází ke světlem indukované separaci náboje, vedoucí k redukci NADP+ a tvorbě pH gradientu využitého pro produkci ATP. Vzhledem k velkému počtu proteinových podjednotek a také kofaktorů včetně Chl, karotenoidů, hemového a nehemového železa, lipidů, iontů atd. patří oba fotosystémy mezi nejsložitější enzymy v přírodě. Díky pokrokům ve strukturní biologii známe jejich detailní struktury (Jordan et al., 2001; Umena et al., 2011), ale méně prozkoumaný je komplikovaný proces jejich biogeneze, vyžadující přísnou regulaci a kontrolu kvality.
Fotosystém II
PSII vzniká postupným sestavením ze čtyř subkomplexů (modulů). Každý z nich se skládá z jedné z velkých Chl-proteinových podjednotek (t.j. klíčových proteinů reakčního centra označovaných D1 a D2, a vnitřních antén CP43 a CP47), přilehlých malých transmembránových podjednotek a pomocných (assembly) faktorů (AF), které usnadňují jednotlivé kroky sestavování (Komenda, Sobotka and Nixon, 2012). AF jsou většinou specifické pro určitý krok biogeneze, a mezi jejich úlohy patří stabilizace protein-proteinových interakcí, ochrana vznikající struktury před předčasnou degradací nebo světelným poškozením a podpora inzerce Chl a dalších kofaktorů. Pro podrobnější vhled do naší práce na toto téma viz např. Komenda et al. (2012), Knoppová et al. (2014), Bečková et al. (2017), Bučinská et al. (2018), Pascual-Aznar et al. (2021), Knoppová et al.(2022). Na kontrole kvality podjednotek PSII, a tedy na "údrbě" funkčního fotosystému, se podílí především tylakoidní sinicové proteázy FtsH, jmenovitě heterokomplex FtsH2/3, který řídí selektivní degradaci podjednotek během tzv. procesu opravy, což je základní mechanismus kompenzující poškození PSII nadměrným světlem (Komenda et al., 2010).
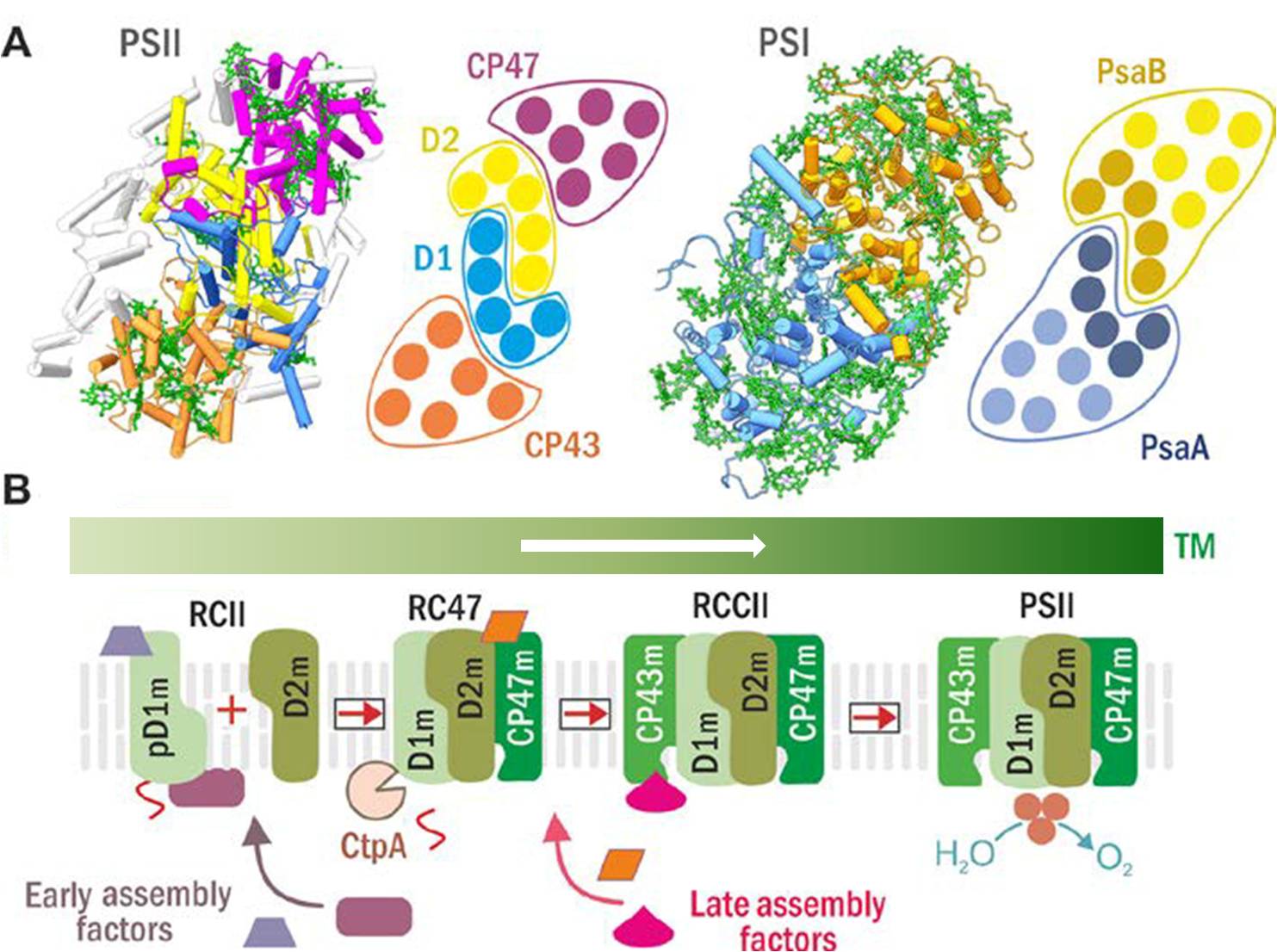
Obr. 1: Struktury PSII a PSI (A) a aktuální model sestavování PSII (B). (A) Struktura PSII (vlevo) a PSI (vpravo) a schematické helikální uspořádání jejich hlavních Chl-proteinů (D1 modře, D2 žlutě, CP47 fialově, CP43 hnědě; PsaA modře se šroubovicemi antény ve světle modré, a PsaB v okrové s anténními šroubovicemi ve světlé okrové). (B) Sestavení PSII probíhá krok po kroku, počínaje syntézou a zabudováním prekurzorového modulu D1 (pD1m) a pokračuje postupným připojením modulů D2, CP47 a CP43. Tvorba komplexu RCII je doprovázena maturací pD1 katalyzované CtpA proteázou. Předpokládá se, že iniciální procesy tvorby PSII jsou prostorově oddělené do specifických domén tylakoidní membrány, tzv. center biogeneze. Jak počáteční, tak pozdější kroky biogeneze PSII jsou facilitovány specifickými pomocnými (assembly) faktory, které nejsou součástí funkčního PSII. (Z návrhu projektu Provázaná biogeneze ..; upraveno.)
Významným představitelem AF fotosystému II je protein Ycf48 (v rostlinách HCF136), který se na počátku biogeneze váže na nově syntetizovaný prekurzor D1 proteinu, klíčové podjednotky PSII, a podporuje účinné spojení s proteinem D2 za vzniku subkomplexu reakčního centra (RCII; Obr. 1B) (Komenda et al., 2008). Kromě toho je Ycf48 potřebný pro efektivní nahrazení poškozeného D1 během opravy PSII (Obr. 3), a existují také důkazy o širší úloze v biogenezi fotosyntetického aparátu. 3D struktura Ycf48 odpovídá sedmilisté vrtuli s konzervovanou argininovou (Arg) doménou na povrchu proteinu, která je důležitá pro vazbu Ycf48 nejen na RCII, ale také na větší komplexy, včetně trimerního fotosystému I (PSI) (Yu et al., 2018). Ve spolupráci s našimi partnery jsme ve vysokém rozlišení vyřešili strukturu RCII tvořenou podjednotkami D1 a D2, PsbI a PsbE/F a Ycf48 (Obr. 2; Zhao et al., 2023). Naše výsledky potvrdily, že se tento pomocný protein váže na lumenální povrch podjednotky D1 prostřednictvím Arg domény, a to přesně v místě, kde se ve funkčním PSII váže kyslík-vyvíjející Mn4CaO5 komplex. To pravděpodobně zabraňuje předčasnému navázání Mn2+ a Ca2+ iontů a tímto způsobem je klíčové vazebné místo chráněno před poškozením způsobeným světlem. Uvolnění Ycf48 z vazby na RCII je pak nezbytné pro světlem indukované sestavení manganového klastru. Model také naznačil interakci mezi Ycf48 a C-terminálním koncem D2, což pomáhá vysvětlit, jak Ycf48 podporuje navázání D1 a D2.
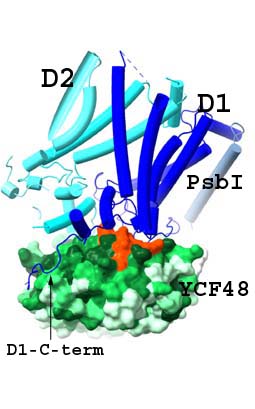 |
Obr. 2: Pomocný faktor Ycf48 (zelená) se váže na klíčovou podjednotku D1 (tmavě modrá) pomocí Arg domény (oranžová) v místě, kde je ve funkčním PSII vázán kyslík-vyvíjející Mn4CaO5 komplex. C-konec D1 proteinu (šipka), který se také účastní ligace manganového komplexu, je odkloněn od tohoto místa vazbou na povrch Ycf48. Toto uspořádání brání předčasnému navázání iontů Mn2+ and Ca2+ a chrání místo před světelným poškozením v průběhu biogeneze. Detailní studie viz Zhao et al. (2023). |
Fotosystém I
Na rozdíl od PSII jsou podrobné informace o biogenezi PSI mnohem omezenější a to se týká zejména rané fáze procesu. V současné době se nám podařilo izolovat hlavní podjednotky heterodimeru reakčního centra, PsaA a PsaB (Obr. 1) s využitím mutantních kmenů, kterým jedna či druhá podjednotka chybí (Gupta a Komenda, v procesu publikování). Izolovaný PsaA obsahuje velké množství Chl, které zhruba odpovídá počtu Chl vázaných na PsaA ve funkčním trimeru PSI (Malavath et al. 2018). Naproti tomu izolovanýPsaB vykazuje pouze velmi omezenou pigmentaci. Přítomnost některých malých membránových podjednotek PSI hovoří pro modulární princip sestavování, který je pravděpodobně společný pro PSI i PSII. Identifikovali jsme také řadu pomocných proteinů specifických pro každou hlavní podjednotku. Kromě již známých assembly faktorů (AF) jsme nalezli také dříve neidentifikované komponenty, potenciálně důležité pro tuto fázi sestavení PSI. Náš současný projekt Raná fáze biogeneze PSI .. je zaměřen na definování podrobného modelu rané fáze sestavování a objasnění role malých podjednotek PSI a AF identifikovaných v časných meziproduktech PSI. Nedávno jsme vyřešili kryo-EM strukturu neúplného PSI z kmene, kterému chybí cytoplazmatická podjednotka PsaC. Zjistili jsme, že malé transmembránové podjednotky (PsaF, I, J, M a K) mohou během procesu sestavování asociovat s heterodimerem PsaA/PsaB bez přítomnosti PsaC a že je tento protein nezbytný pro vazbu dalších dvou cytoplazmatických podjednotek. Dále přispívá také k navázání PsaL, který silně interaguje s PsaD a usnadňuje trimerizaci PSI. PsaC je rovněž důležitý pro správnou konfiguraci vazebného místa pro železo-sírný klastr FX (Ferroussier, Gupta et al., 2025).
Provázaná biogeneze PSI a PSII
Asociace specifických meziproduktů PSII s monomerním či trimerním PSI (Bečková et al., 2017; Strašková et al. 2018; Kiss et al., 2019; Knoppová et al., 2022) nebo s enzymy chlorofylové dráhy (Dobáková et al. al., 2009 a nepublikované výsledky), stejně jako spojení Chl-syntázy, konečného enzymu biosyntézy Chl, s YidC inzertázou a Sec translokázou (Chidgey et al. 2014) naznačují, že tyto dva fotosystémy pravděpodobně sdílejí společný buněčný aparát integrující procesy vedoucí k tvorbě a zabudování Chl-proteinů se syntézou Chl (viz Provázaná biogeneze ...). Předpokládáme existenci funkčních domén řídících nasměrování de novo vzniklých molekul Chl k podjednotkám PSI či PSII, tak jak se mění poměr PSI:PSII za různých podmínek prostředí, a také se domníváme, že navázání meziproduktů biogeneze fotosystému II na fotosystém I chrání vznikající PSII před světelným poškozením díky "přelévání" (spillover) absorbované energie směrem na PSI.
Proteázy FtsH - úloha ve fotosyntéze a dalších buněčných procesech
FtsH proteázy patří mezi intramembránové ATP-dependentní metaloproteázy, které nacházíme v prokaryotech, mitochondriích a chloroplastech. Většina sinic, včetně Synechocystis, má čtyři homology FtsH (FtsH1 až FtsH4). Sestavují se do tří oligomerních komplexů: dva heterooligomery FtsH1/3 a FtsH2/3 a jeden homooligomer FtsH4. Komplex FtsH1/3 se nachází v plazmatické membráně, zatímco hojnější FtsH2/3 a FtsH4 jsou lokalizovány v tylakoidních membránách. Bylo prokázáno, že FtsH1/3 u Synechocystis je rozhodující pro jemné doladění transkripční odpovědi na živinový stres. Na druhé straně, tylakoidní FtsH2/3 řídí selektivní degradaci podjednotek PSII během procesu opravy, což je základní mechanismus kompenzující poškození PSII nadměrným světlem, a tak hraje klíčovou roli při kontrole kvality komplexů PSII. Delece genu ftsH2 vede k abnormální citlivosti na světlo a k fotoinhibici PSII, ale také má za důsledek snížení hladiny PSI a obsahu buněčného Chl. Druhý tylakoidní komplex FtsH4 se podílí na biogenezi PSII a PSI a aklimatizaci na náhlé změny intenzity světla regulací hladiny proteinů indukovaných vysokým světlem (Hlips) zapojených do fotoprotekce vznikajících komplexů PSII. Pro recentní review viz Krynická and Komenda (2024).
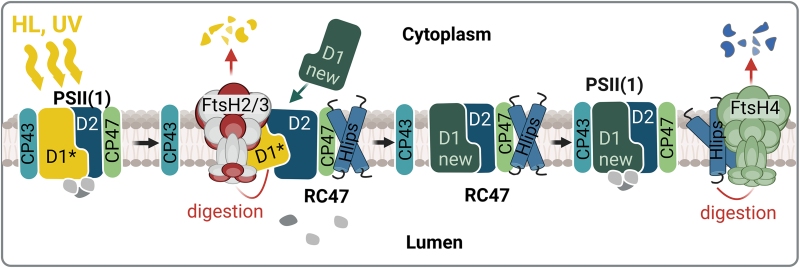
Obr. 3: Úloha tylakoidních FtsH ve fotoprotekci komplexu PSII. Při stresu způsobeném nadměrným viditelným světlem či UV je rychlá inaktivace PSII především důsledkem poškození D1 proteinu (D1*, žlutá), což vede k destabilizaci vazby CP43 (modrozelená) v PSII a akumulaci intermediátu RC47. To umožňuje přístup FtsH2/3 k poškozenému D1, který je pak rychle degradován a nahrazen nově syntetizovanou D1 (new D1, tmavě zelená)). Následně se anténa CP43 znovu připojí k opravenému RC47. Procesu opravy se účastní světlem vysoce indukované proteiny (Hlips, modrá), které chrání meziprodukty PSII před oxidativním poškozením tím, že zhášejí energii excitovaného Chl. Po dokončení opravy PSII se Hlips uvolní z komplexu a jejich hladina je regulována proteázou FtsH4 (Krynická and Komenda, 2024).
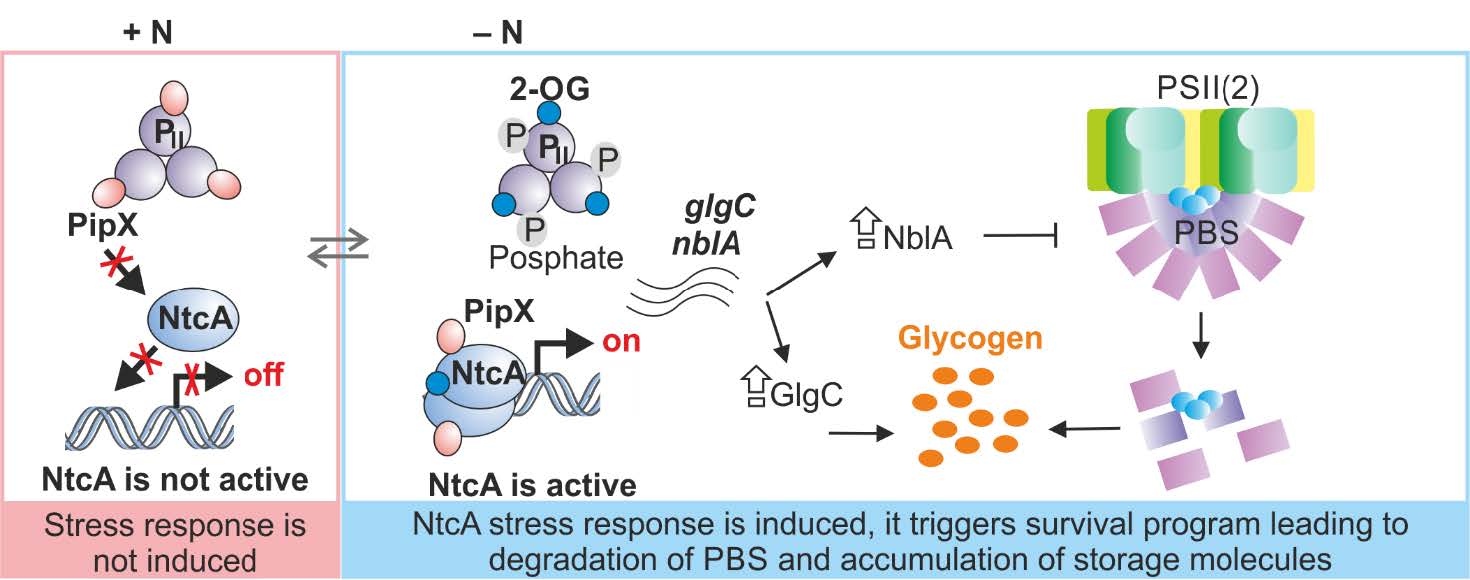
Modifikace NtcA regulonu a možnosti jeho využití v biotechnologii
Globální regulační dráhy v sinicích, jako je NtcA, jsou slibnými cíli pro zelenou biotechnologii. Dráha NtcA představuje integrovaný metabolický program, který reaguje na nedostatek dusíku, což vede k akumulaci zásobních polymerů. Nedávný výzkum odhalil klíčovou roli proteolytického komplexu FtsH1/3 v aktivaci transkripční odpovědi NtcA na omezení dusíku. Přesný mechanismus této aktivace však zůstává neznámý. V tomto projektu chceme u sinice Synechocystis indukovat odpověď NtcA za nestresových podmínek, abychom zabránili inhibici fotosyntézy a tím zvýšili akumulaci zásobních sacharidů. Toho se snažíme dosáhnout zkonstruováním faktoru typu NtcA, který se exprimuje i za podmínek dostatku dusíku. Tento přístup doplníme také modulací hladiny komplexu FtsH1/3 pro zvýšení odpovědi. Dopad přeprogramování NtcA regulonu na celkový buněčný metabolismus, včetně syntézy zásobních sloučenin, vyhodnocujeme pomocí nejmodernějších metod, jako jsou omické analýzy.